Optogenetics
Optogenetics (from Greek, Modern optikós, meaning 'seen, visible') is a biological technique that involves the use of light to control cells in living tissue, typically neurons, that have been genetically modified to express light-sensitive ion channels. It is a neuromodulation method that uses a combination of techniques from optics and genetics to control and monitor the activities of individual neurons in living tissue—even within freely-moving animals—and to precisely measure these manipulation effects in real-time.[1] The key reagents used in optogenetics are light-sensitive proteins. Neuronal control is achieved using optogenetic actuators like channelrhodopsin, halorhodopsin, and archaerhodopsin, while optical recording of neuronal activities can be made with the help of optogenetic sensors for calcium (GCaMP), vesicular release (synapto-pHluorin), neurotransmitter (GluSnFRs), or membrane voltage (arc lightning, ASAP1).[2][3] Control (or recording) of activity is restricted to genetically defined neurons and performed in a spatiotemporal-specific manner by light.
In 2010, optogenetics was chosen as the "Method of the Year" across all fields of science and engineering by the interdisciplinary research journal Nature Methods.[4] At the same time, optogenetics was highlighted in the article on "Breakthroughs of the Decade" in the academic research journal Science.[5] These journals also referenced recent public-access general-interest video Method of the year video and textual SciAm summaries of optogenetics.
Contents
History[edit]
The "far-fetched" possibility of using light for selectively controlling precise neural activity (action potential) patterns within subtypes of cells in the brain was thought of by Francis Crick in his Kuffler Lectures at the University of California in San Diego in 1999.[6] An earlier use of light to activate neurons was carried out by Richard Fork,[7] who demonstrated laser activation of neurons within intact tissue, although not in a genetically-targeted manner. The earliest genetically targeted method that used light to control rhodopsin-sensitized neurons was reported in January 2002, by Boris Zemelman and Gero Miesenböck, who employed Drosophila rhodopsin cultured mammalian neurons.[8] In 2003, Zemelman and Miesenböck developed a second method for light-dependent activation of neurons in which single inotropic channels TRPV1, TRPM8 and P2X2 were gated by photocaged ligands in response to light.[9] Beginning in 2004, the Kramer and Isacoff groups developed organic photoswitches or "reversibly caged" compounds in collaboration with the Trauner group that could interact with genetically introduced ion channels.[10][11] TRPV1 methodology, albeit without the illumination trigger, was subsequently used by several laboratories to alter feeding, locomotion and behavioral resilience in laboratory animals.[12][13][14] However, light-based approaches for altering neuronal activity were not applied outside the original laboratories, likely because the easier to employ channelrhodopsin was cloned soon thereafter.[15]
Peter Hegemann, studying the light response of green algae at the University of Regensburg, had discovered photocurrents that were too fast to be explained by the classic g-protein-coupled animal rhodopsins.[16] Teaming up with the electrophysiologist Georg Nagel at the Max Planck Institute in Frankfurt, they could demonstrate that a single gene from the alga Chlamydomonas produced large photocurents when expressed in the oocyte of a frog.[17] To identify expressing cells, they replaced the cytoplasmic tail of the algal protein with the fluorescent protein YFP, generating the first generally applicable optogenetic tool.[15]
Zhuo-Hua Pan of Wayne State University, researching on restore sight to blindness, thought about using channelrhodopsin when it came out in late 2003. By February 2004, he was trying channelrhodopsin out in ganglion cells—the neurons in our eyes that connect directly to the brain—that he had cultured in a dish. Indeed, the transfected neurons became electrically active in response to light. In 2005, Zhuo-Hua Pan reported successful in-vivo transfection of channelrhodopsin in retinal ganglion cells of mice, and electrical responses to photostimulation in retinal slice culture[18]
In April 2005, Susana Lima and Miesenböck reported the first use of genetically-targeted P2X2 photostimulation to control the behaviour of an animal.[19] They showed that photostimulation of genetically circumscribed groups of neurons, such as those of the dopaminergic system, elicited characteristic behavioural changes in fruit flies.
In August 2005, Karl Deisseroth's laboratory in the Bioengineering Department at Stanford including graduate students Ed Boyden and Feng Zhang published the first demonstration of a single-component optogenetic system in cultured mammalian neurons,[20][21] using the channelrhodopsin-2(H134R)-eYFP construct from Nagel and Hegemann.[15]
The groups of Gottschalk and Nagel were first to use channelrhodopsin-2 for controlling neuronal activity in an intact animal, showing that motor patterns in the roundworm Caenorhabditis elegans could be evoked by light stimulation of genetically selected neural circuits (published in December 2005).[22] In mice, controlled expression of optogenetic tools is often achieved with cell-type-specific Cre/loxP methods developed for neuroscience by Joe Z. Tsien back in the 1990s[23] to activate or inhibit specific brain regions and cell-types in vivo.[24]
The primary tools for optogenetic recordings have been genetically encoded calcium indicators (GECIs). The first GECI to be used to image activity in an animal was cameleon, designed by Atsushi Miyawaki, Roger Tsien and coworkers.[25] Cameleon was first used successfully in an animal by Rex Kerr, William Schafer and coworkers to record from neurons and muscle cells of the nematode C. elegans.[26] Cameleon was subsequently used to record neural activity in flies[27] and zebrafish.[28] In mammals, the first GECI to be used in vivo was GCaMP,[29] first developed by Nakai and coworkers.[30] GCaMP has undergone numerous improvements, and GCaMP6[31] in particular has become widely used throughout neuroscience.
In 2010, Karl Deisseroth at Stanford University was awarded the inaugural HFSP Nakasone Award "for his pioneering work on the development of optogenetic methods for studying the function of neuronal networks underlying behavior". In 2012, Gero Miesenböck was awarded the InBev-Baillet Latour International Health Prize for "pioneering optogenetic approaches to manipulate neuronal activity and to control animal behaviour." In 2013, Ernst Bamberg, Ed Boyden, Karl Deisseroth, Peter Hegemann, Gero Miesenböck and Georg Nagel were awarded The Brain Prize for "their invention and refinement of optogenetics."[32][33] Karl Deisseroth was awarded the Else Kröner Fresenius Research Prize 2017 (4 million euro) for his "contributions to the understanding of the biological basis of psychiatric disorders".
Description[edit]


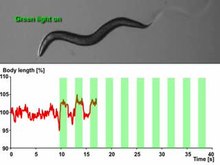

Optogenetics provides millisecond-scale temporal precision which allows the experimenter to keep pace with fast biological information processing (for example, in probing the causal role of specific action potential patterns in defined neurons). Indeed, to probe the neural code, optogenetics by definition must operate on the millisecond timescale to allow addition or deletion of precise activity patterns within specific cells in the brains of intact animals, including mammals (see Figure 1). By comparison, the temporal precision of traditional genetic manipulations (employed to probe the causal role of specific genes within cells, via "loss-of-function" or "gain of function" changes in these genes) is rather slow, from hours or days to months. It is important to also have fast readouts in optogenetics that can keep pace with the optical control. This can be done with electrical recordings ("optrodes") or with reporter proteins that are biosensors, where scientists have fused fluorescent proteins to detector proteins. An example of this is voltage-sensitive fluorescent protein (VSFP2).[37] Additionally, beyond its scientific impact optogenetics represents an important case study in the value of both ecological conservation (as many of the key tools of optogenetics arise from microbial organisms occupying specialized environmental niches), and in the importance of pure basic science as these opsins were studied over decades for their own sake by biophysicists and microbiologists, without involving consideration of their potential value in delivering insights into neuroscience and neuropsychiatric disease.[38]
Light-activated proteins: channels, pumps and enzymes
The hallmark of optogenetics therefore is introduction of fast light-activated channels, pumps, and enzymes that allow temporally precise manipulation of electrical and biochemical events while maintaining cell-type resolution through the use of specific targeting mechanisms. Among the microbial opsins which can be used to investigate the function of neural systems are the channelrhodopsins (ChR2, ChR1, VChR1, and SFOs) to excite neurons and anion-conducting channelrhodopsins for light-induced inhibition. Indirectly light-controlled potassium channels have recently been engineered to prevent action potential generation in neurons during blue light illumination.[39][40] Light-driven ion pumps are also used to inhibit neuronal activity, e.g. halorhodopsin (NpHR),[41] enhanced halorhodopsins (eNpHR2.0 and eNpHR3.0, see Figure 2),[42] archaerhodopsin (Arch), fungal opsins (Mac) and enhanced bacteriorhodopsin (eBR).[43]
Optogenetic control of well-defined biochemical events within behaving mammals is now also possible. Building on prior work fusing vertebrate opsins to specific G-protein coupled receptors[44] a family of chimeric single-component optogenetic tools was created that allowed researchers to manipulate within behaving mammals the concentration of defined intracellular messengers such as cAMP and IP3 in targeted cells.[45] Other biochemical approaches to optogenetics (crucially, with tools that displayed low activity in the dark) followed soon thereafter, when optical control over small GTPases and adenylyl cyclases was achieved in cultured cells using novel strategies from several different laboratories.[46][47][48][49][50] This emerging repertoire of optogenetic probes now allows cell-type-specific and temporally precise control of multiple axes of cellular function within intact animals.[51]
Hardware for light application
Another necessary factor is hardware (e.g. integrated fiberoptic and solid-state light sources) to allow specific cell types, even deep within the brain, to be controlled in freely behaving animals. Most commonly, the latter is now achieved using the fiberoptic-coupled diode technology introduced in 2007,[52][53][54] though to avoid use of implanted electrodes, researchers have engineered ways to inscribe a "window" made of zirconia that has been modified to be transparent and implanted in mice skulls, to allow optical waves to penetrate more deeply to stimulate or inhibit individual neurons.[55] To stimulate superficial brain areas such as the cerebral cortex, optical fibers or LEDs can be directly mounted to the skull of the animal. More deeply implanted optical fibers have been used to deliver light to deeper brain areas. Complementary to fiber-tethered approaches, completely wireless techniques have been developed utilizing wirelessly delivered power to headborne LEDs for unhindered study of complex behaviors in freely behaving organisms.[56]
Expression of optogenetic actuators
Optogenetics also necessarily includes the development of genetic targeting strategies such as cell-specific promoters or other customized conditionally-active viruses, to deliver the light-sensitive probes to specific populations of neurons in the brain of living animals (e.g. worms, fruit flies, mice, rats, and monkeys). In invertebrates such as worms and fruit flies some amount of all-trans-retinal (ATR) is supplemented with food. A key advantage of microbial opsins as noted above is that they are fully functional without the addition of exogenous co-factors in vertebrates.[54]
Technique[edit]

The technique of using optogenetics is flexible and adaptable to the experimenter's needs. For starters, experimenters genetically engineer a microbial opsin based on the gating properties (rate of excitability, refractory period, etc..) required for the experiment.
There is a challenge in introducing the microbial opsin, an optogenetic actuator, into a specific region of the organism in question. A rudimentary approach is to introduce an engineered viral vector that contains the optogenetic actuator gene attached to a recognizable promoter such as CAMKIIα. This allows for some level of specificity as cells that already contain and can translate the given promoter will be infected with the viral vector and hopefully express the optogenetic actuator gene.
Another approach is the creation of transgenic mice where the optogenetic actuator gene is introduced into mice zygotes with a given promoter, most commonly Thy1. Introduction of the optogenetic actuator at an early stage allows for a larger genetic code to be incorporated and as a result, increases the specificity of cells to be infected.
A third and rather novel approach that has been developed is creating transgenic mice with Cre recombinase, an enzyme that catalyzes recombination between two lox-P sites. Then by introducing an engineered viral vector containing the optogenetic actuator gene in between two lox-P sites, only the cells containing the Cre recombinase will express the microbial opsin. This last technique has allowed for multiple modified optogenetic actuators to be used without the need to create a whole line of transgenic animals every time a new microbial opsin is needed.
After the introduction and expression of the microbial opsin, depending on the type of analysis being performed, application of light can be placed at the terminal ends or the main region where the infected cells are situated. Light stimulation can be performed with a vast array of instruments from light emitting diodes (LEDs) or diode-pumped solid state (DPSS). These light sources are most commonly connected to a computer through a fiber optic cable. Recent advances include the advent of wireless head-mounted devices that also apply LED to targeted areas and as a result give the animal more freedom of mobility to reproduce in vivo results.[58][59]
Issues[edit]
Although already a powerful scientific tool, optogenetics, according to Doug Tischer & Orion D. Weiner of the University of California San Francisco, should be regarded as a "first-generation GFP" because of its immense potential for both utilization and optimization.[60] With that being said, the current approach to optogenetics is limited primarily by its versatility. Even within the field of Neuroscience where it is most potent, the technique is less robust on a subcellular level.[61]
Selective expression[edit]
One of the main problems of optogenetics is that not all the cells in question may express the microbial opsin gene at the same level. Thus, even illumination with a defined light intensity will have variable effects on individual cells. Optogenetic stimulation of neurons in the brain is even less controlled as the light intensity drops exponentially from the light source (e.g. implanted optical fiber).
Moreover, mathematical modelling shows that selective expression of opsin in specific cell types can dramatically alter the dynamical behavior of the neural circuitry. In particular, optogenetic stimulation that preferentially targets inhibitory cells can transform the excitability of the neural tissue from Type 1 — where neurons operate as integrators — to Type 2 where neurons operate as resonators.[62] Type 1 excitable media sustain propagating waves of activity whereas Type 2 excitable media do not. The transformation from one to the other explains how constant optical stimulation of primate motor cortex elicits gamma-band (40–80 Hz) oscillations in the manner of a Type 2 excitable medium. Yet those same oscillations propagate far into the surrounding tissue in the manner of a Type 1 excitable medium.[63]
Nonetheless, it remains difficult to target opsin to defined subcellular compartments, e.g. the plasma membrane, synaptic vesicles, or mitochondria.[61][64] Restricting the opsin to specific regions of the plasma membrane such as dendrites, somata or axon terminals would provide a more robust understanding of neuronal circuitry.[61]
Kinetics and synchronization[edit]
An issue with channelrhodopsin-2 is that its gating properties don't mimic in vivo cation channels of cortical neurons. A solution to this issue with a protein's kinetic property is introduction of variants of channelrhodopsin-2 with more favorable kinetics.[55][56]
Another one of the technique's limitations is that light stimulation produces a synchronous activation of infected cells and this removes any individual cell properties of activation among the population affected. Therefore, it is difficult to understand how the cells in the population affected communicate with one another or how their phasic properties of activation may relate to the circuitry being observed.
Optogenetic activation has been combined with functional magnetic resonance imaging (ofMRI) to elucidate the connectome, a thorough map of the brain’s neural connections. The results, however, are limited by the general properties of fMRI.[61][65] The readouts from this neuroimaging procedure lack the spatial and temporal resolution appropriate for studying the densely packed and rapid-firing neuronal circuits.[65]
Excitation spectrum[edit]
The opsin proteins currently in use have absorption peaks across the visual spectrum, but remain considerable sensitivity to blue light.[61] This spectral overlap makes it very difficult to combine opsin activation with genenetically encoded indictors (GEVIs, GECIs, GluSnFR, synapto-pHluorin), most of which need blue light excitation. Opsins with infrared activation would, at a standard irradiance value, increase light penetration and augment resolution through reduction of light scattering.
Additional data indicates that the absorption spectra of organic dyes and fluorescent proteins, used in optogenetics applications, extends from around 250 nm to around 600 nm. Particular organic compounds used in discrete portions of this range include: retinals, flavins, folates, p-coumaric acids, phytochrome chromophotes, cobalamins, and at least six fluorescent proteins including mOrange and mCherry.[66]
Applications[edit]
The field of optogenetics has furthered the fundamental scientific understanding of how specific cell types contribute to the function of biological tissues such as neural circuits in vivo (see references from the scientific literature below). Moreover, on the clinical side, optogenetics-driven research has led to insights into Parkinson's disease[67][68] and other neurological and psychiatric disorders. Indeed, optogenetics papers in 2009 have also provided insight into neural codes relevant to autism, Schizophrenia, drug abuse, anxiety, and depression.[43][69][70][71]
Identification of particular neurons and networks[edit]
Amygdala[edit]
Optogenetic approaches have been used to map neural circuits in the amygdala that contribute to fear conditioning.[72][73][74][75] One such example of a neural circuit is the connection made from the basolateral amygdala to the dorsal-medial prefrontal cortex where neuronal oscillations of 4 Hz have been observed in correlation to fear induced freezing behaviors in mice. Transgenic mice were introduced with channelrhodoposin-2 attached with a parvalbumin-Cre promoter that selectively infected interneurons located both in the basolateral amygdala and the dorsal-medial prefrontal cortex responsible for the 4 Hz oscillations. The interneurons were optically stimulated generating a freezing behavior and as a result provided evidence that these 4 Hz oscillations may be responsible for the basic fear response produced by the neuronal populations along the dorsal-medial prefrontal cortex and basolateral amygdala.[76]
Olfactory bulb[edit]
Optogenetic activation of olfactory sensory neurons was critical for demonstrating timing in odor processing[77] and for mechanism of neuromodulatory mediated olfactory guided behaviors (e.g. aggression, mating)[78] In addition, with the aid of optogenetics, evidence has been reproduced to show that the "afterimage" of odors is concentrated more centrally around the olfactory bulb rather than on the periphery where the olfactory receptor neurons would be located. Transgenic mice infected with channel-rhodopsin Thy1-ChR2, were stimulated with a 473 nm laser transcranially positioned over the dorsal section of the olfactory bulb. Longer photostimulation of mitral cells in the olfactory bulb led to observations of longer lasting neuronal activity in the region after the photostimulation had ceased, meaning the olfactory sensory system is able to undergo long term changes and recognize differences between old and new odors.[79]
Nucleus accumbens[edit]
Optogenetics, freely moving mammalian behavior, in vivo electrophysiology, and slice physiology have been integrated to probe the cholinergic interneurons of the nucleus accumbens by direct excitation or inhibition. Despite representing less than 1% of the total population of accumbal neurons, these cholinergic cells are able to control the activity of the dopaminergic terminals that innervate medium spiny neurons (MSNs) in the nucleus accumbens.[80] These accumbal MSNs are known to be involved in the neural pathway through which cocaine exerts its effects, because decreasing cocaine-induced changes in the activity of these neurons has been shown to inhibit cocaine conditioning. The few cholinergic neurons present in the nucleus accumbens may prove viable targets for pharmacotherapy in the treatment of cocaine dependence[43]

Prefrontal cortex[edit]
In vivo and in vitro recordings (by the Cooper laboratory) of individual CAMKII AAV-ChR2 expressing pyramidal neurons within the prefrontal cortex demonstrated high fidelity action potential output with short pulses of blue light at 20 Hz (Figure 1).[34] The same group recorded complete green light-induced silencing of spontaneous activity in the same prefrontal cortical neuronal population expressing an AAV-NpHR vector (Figure 2).[34]
Heart[edit]
Optogenetics was applied on atrial cardiomyocytes to end spiral wave arrhythmias, found to occur in atrial fibrillation, with light.[81] This method is still in the development stage. A recent study explored the possibilities of optogenetics as a method to correct for arrythmias and resynchronize cardiac pacing. The study introduced channelrhodopsin-2 into cardiomyocytes in ventricular areas of hearts of transgenic mice and performed in vitro studies of photostimulation on both open-cavity and closed-cavity mice. Photostimulation led to increased activation of cells and thus increased ventricular contractions resulting in increasing heart rates. In addition, this approach has been applied in cardiac resynchronization therapy (CRT) as a new biological pacemaker as a substitute for electrode based-CRT.[82] Lately, optogenetics has been used in the heart to defibrillate ventricular arrhythmias with local epicardial illumination,[83] a generalized whole heart illumination[84] or with customized stimulation patterns based on arrhythmogenic mechanisms in order to lower defibrillation energy.[85]
Spiral ganglion[edit]
Optogenetic stimulation of the spiral ganglion in deaf mice restored auditory activity.[86][87] Optogenetic application onto the cochlear region allows for the stimulation or inhibition of the spiral ganglion cells (SGN). In addition, due to the characteristics of the resting potentials of SGN's, different variants of the protein channelrhodopsin-2 have been employed such as Chronos and CatCh. Chronos and CatCh variants are particularly useful in that they have less time spent in their deactivated states, which allow for more activity with less bursts of blue light emitted. The result being that the LED producing the light would require less energy and the idea of cochlear prosthetics in association with photo-stimulation, would be more feasible.[88]
Brainstem[edit]
Optogenetic stimulation of a modified red-light excitable channelrhodopsin (ReaChR) expressed in the facial motor nucleus enabled minimally invasive activation of motoneurons effective in driving whisker movements in mice.[89] One novel study employed optogenetics on the Dorsal Ralphe Nucleus to both activate and inhibit dopaminergic release onto the ventral tegmental area. To produce activation transgenic mice were infected with channelrhodopsin-2 with a TH-Cre promoter and to produce inhibition the hyperpolarizing opsin NpHR was added onto the TH-Cre promoter. Results showed that optically activating dopaminergic neurons led to an increase in social interactions, and their inhibition decreased the need to socialize only after a period of isolation.[90]
Precise temporal control of interventions[edit]
The currently available optogenetic actuators allow for the accurate temporal control of the required intervention (i.e. inhibition or excitation of the target neurons) with precision routinely going down to the millisecond level. Therefore, experiments can now be devised where the light used for the intervention is triggered by a particular element of behavior (to inhibit the behavior), a particular unconditioned stimulus (to associate something to that stimulus) or a particular oscillatory event in the brain (to inhibit the event). This kind of approach has already been used in several brain regions:
Hippocampus[edit]
Sharp waves and ripple complexes (SWRs) are distinct high frequency oscillatory events in the hippocampus thought to play a role in memory formation and consolidation. These events can be readily detected by following the oscillatory cycles of the on-line recorded local field potential. In this way the onset of the event can be used as a trigger signal for a light flash that is guided back into the hippocampus to inhibit neurons specifically during the SWRs and also to optogenetically inhibit the oscillation itself[91] These kinds of "closed-loop" experiments are useful to study SWR complexes and their role in memory.
Cellular biology/cell signaling pathways[edit]
The optogenetic toolkit has proven pivotal for the field of neuroscience as it allows precise manipulation of neuronal excitability. Moreover, this technique has been shown to extend outside neurons to an increasing number of proteins and cellular functions.[60] Cellular scale modifications including manipulation of contractile forces relevant to cell migration, cell division and wound healing have been optogenetically manipulated.[92] The field has not developed to the point where processes crucial to cellular and developmental biology and cell signaling including protein localization, post-translational modification and GTP loading can be consistently controlled via optogenetics.[60]
Photosensitive proteins utilized in various cell signaling pathways[edit]
While this extension of optogenetics remains to be further investigated, there are various conceptual methodologies that may prove to immediately robust. There is a considerable body of literature outlining photosensitive proteins that have been utilized in cell signaling pathways.[60] CRY2, LOV, DRONPA and PHYB are photosynthetic proteins involved in inducible protein association whereby activation via light can induce/turn off a signaling cascade via recruitment of a signaling domain to its respective substrate.[93][94][95][96] LOV and PHYB are photosensitive proteins that engage in homodimerization and/or heterodimerization to recruit some DNA-modifying protein, translocate to the site of DNA and alter gene expression levels.[97][98][99] CRY2, a protein that inherently clusters when active, has been fused with signaling domains and subsequently photoactivated allowing for clustering-based activation.[100] Proteins LOV and Dronpa have also been adapted to cell signaling manipulation; exposure to light induces conformational changes in the photosensitive protein which can subsequently reveal a previously obscured signaling domain and/or activate a protein that was otherwise allosterically inhibited.[101][102] LOV has been fused to caspase 3 to produce a construct capable of inducing apoptosis upon light stimulation.[103]
Optogenetic temporal control of signals[edit]
A different set of signaling cascades respond to stimulus timing duration and dynamics.[104] Adaptive signaling pathways, for instance, adjust in accordance to the current level of the projected stimulus and display activity only when these levels change as opposed to responding to absolute levels of the input.[105] Stimulus dynamics also can trigger activity; treating PC12 cells with epidermal growth factor (inducing a transient profile of ERK activity) leads to cellular proliferation whereas introduction of nerve growth factor (inducing a sustained profile of ERK activity) is associated with a different cellular decision whereby the PC12 cells differentiate into neuron-like cells.[106] This discovery was guided pharmacologically but the finding was replicated utilizing optogenetic inputs instead.[107] This ability to optogenetically control signals for various time durations is being explored to elucidate various cell signaling pathways where there is not a strong enough understanding to utilize either drug/genetic manipulation.[60]
References[edit]
- ^ Deisseroth, K.; Feng, G.; Majewska, A. K.; Miesenbock, G.; Ting, A.; Schnitzer, M. J. (2006). "Next-Generation Optical Technologies for Illuminating Genetically Targeted Brain Circuits". Journal of Neuroscience. 26 (41): 10380–6. doi:10.1523/JNEUROSCI.3863-06.2006. PMC 2820367. PMID 17035522.
- ^ Mancuso, J. J.; Kim, J.; Lee, S.; Tsuda, S.; Chow, N. B. H.; Augustine, G. J. (2010). "Optogenetic probing of functional brain circuitry". Experimental Physiology. 96 (1): 26–33. doi:10.1113/expphysiol.2010.055731. PMID 21056968.
- ^ Treger, Jeremy (2015). "Single-molecule fluorimetry and gating currents inspire an improved optical voltage indicator". eLife. 4: e10482. doi:10.7554/eLife.10482. PMC 4658195. PMID 26599732.
- ^ Primer on Optogenetics: Pastrana, Erika (2010). "Optogenetics: Controlling cell function with light". Nature Methods. 8 (1): 24–25. doi:10.1038/nmeth.f.323.
Editorial: "Method of the Year 2010". Nature Methods. 8 (1): 1. 2010. doi:10.1038/nmeth.f.321.
Commentary: Deisseroth, Karl (2010). "Optogenetics". Nature Methods. 8 (1): 26–9. doi:10.1038/nmeth.f.324. PMID 21191368. - ^ News, Staff (2010). "Insights of the decade. Stepping away from the trees for a look at the forest. Introduction". Science. 330 (6011): 1612–3. Bibcode:2010Sci...330.1612.. doi:10.1126/science.330.6011.1612. PMID 21163985.
- ^ Crick, F. (December 1999). "The impact of molecular biology on neuroscience". Philosophical Transactions of the Royal Society B. 354 (1392): 2021–25. doi:10.1098/rstb.1999.0541. PMC 1692710. PMID 10670022.
- ^ Fork, R. L. (March 1971). "Laser stimulation of nerve cells in Aplysia". Science. 171 (3974): 907–8. Bibcode:1971Sci...171..907F. doi:10.1126/science.171.3974.907. PMID 5541653.
- ^ Zemelman, B. V.; Lee, G. A.; Ng, M.; Miesenböck, G. (2002). "Selective photostimulation of genetically chARGed neurons". Neuron. 33 (1): 15–22. doi:10.1016/S0896-6273(01)00574-8. PMID 11779476.
- ^ Zemelman, B. V.; Nesnas, N.; Lee, G.A.; Miesenböck, G. (2003). "Photochemical gating of heterologous ion channels: Remote control over genetically designated populations of neurons". PNAS. 100: 1352–7. Bibcode:2003PNAS..100.1352Z. doi:10.1073/pnas.242738899. PMC 298776. PMID 12540832.
- ^ Banghart, M; Borges, K; Isacoff, E; Trauner, R. H. (21 November 2004). "Light-activated ion channels for remote control of neuronal firing". Nature Neuroscience. 7 (12): 1381–1386. doi:10.1038/nn1356. PMC 1447674. PMID 15558062.
- ^ Volgraf,, M.; Gorostiza, P.; Numano, R.; Kramer, R. H.; Isacoff, E. Y. (11 December 2005). "Allosteric control of an ionotropic glutamate receptor with an optical switch". Nature Chemical Biology. 2 (1): 47–52. doi:10.1038/nchembio756. PMC 1447676. PMID 16408092.
- ^ Arenkiel, BR; Klein, ME; Davison, IG; Katz, LC; Ehlers, MD (2008). "Genetic control of neuronal activity in mice conditionally expressing TRPV1". Nature Methods. 5: 299–302. doi:10.1038/nmeth.1190. PMC 3127246. PMID 18327266. Retrieved 2017-03-15.
- ^ "Transient activation of specific neurons in mice by selective expression of the capsaicin receptor". Bibcode:2012NatCo...3E.746G. doi:10.1038/ncomms1749. Retrieved 2017-03-15.
- ^ "Synaptic Modifications in the Medial Prefrontal Cortex in Susceptibility and Resilience to Stress". JNeurosci. Retrieved 2017-03-15.
- ^ a b c Nagel,, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. (25 November 2003). "Channelrhodopsin-2, a directly light-gated cation-selective membrane channel". Proc Natl Acad Sci U S A. 100 (24): 13940–5. Bibcode:2003PNAS..10013940N. doi:10.1073/pnas.1936192100. PMC 283525. PMID 14615590.
- ^ Harz, Hartmann; Hegemann, Peter (1991-06-06). "Rhodopsin-regulated calcium currents in Chlamydomonas". Nature. 351 (6326): 489–491. Bibcode:1991Natur.351..489H. doi:10.1038/351489a0.
- ^ Nagel, Georg; Ollig, Doris; Fuhrmann, Markus; Kateriya, Suneel; Musti, Anna Maria; Bamberg, Ernst; Hegemann, Peter (2002-06-28). "Channelrhodopsin-1: A Light-Gated Proton Channel in Green Algae". Science. 296 (5577): 2395–2398. Bibcode:2002Sci...296.2395N. doi:10.1126/science.1072068. ISSN 0036-8075. PMID 12089443.
- ^ Bi, Anding; Cui, Jinjuan; Ma, Yu-Ping; Olshevskaya, Elena; Pu, Mingliang; Dizhoor, Alexander M.; Pan, Zhuo-Hua (2006). "Ectopic Expression of a Microbial-Type Rhodopsin Restores Visual Responses in Mice with Photoreceptor Degeneration". Neuron. 50 (1): 23–33. doi:10.1016/j.neuron.2006.02.026. ISSN 0896-6273. PMC 1459045. PMID 16600853.
- ^ Lima, S. Q.; Miesenböck, G. (2005). "Remote Control of Behavior through Genetically Targeted Photostimulation of Neurons". Cell. 121 (1): 141–152. doi:10.1016/j.cell.2005.02.004. PMID 15820685.
- ^ Boyden, E. S.; Zhang, F.; Bamberg, E.; Nagel, G.; Deisseroth, K. (2005). "Millisecond-timescale, genetically targeted optical control of neural activity". Nat. Neurosci. 8 (9): 1263–8. doi:10.1038/nn1525. PMID 16116447.
- ^ Li,, X.; Gutierrez, D. V.; Hanson, M. G.; Han, J.; Mark, M. D.; Chiel, H.; Hegemann, P.; Landmesser, L. T.; Herlitze, S. (14 October 2005). "Fast noninvasive activation and inhibition of neural and network activity by vertebrate rhodopsin and green algae channelrhodopsin". Proc Natl Acad Sci U S A. 102 (49): 17816–21. Bibcode:2005PNAS..10217816L. doi:10.1073/pnas.0509030102. PMC 1292990. PMID 16306259.
- ^ Nagel, G.; Brauner, M.; Liewald, J. F.; Adeishvili, N.; Bamberg, E.; Gottschalk, A. (December 2005). "Light activation of channelrhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses". Curr. Biol. 15 (24): 2279–84. doi:10.1016/j.cub.2005.11.032. PMID 16360690.
- ^ Tsien, JZ; et al. (Dec 1996). "Subregion- and cell type-restricted gene knockout in mouse brain". Cell. 87 (7): 1317–26. doi:10.1016/S0092-8674(00)81826-7. PMID 8980237.
- ^ Tsien, JZ (2016). "Cre-Lox Neurogenetics: 20 Years of Versatile Applications in Brain Research and Counting". Front Genet. 7: 19. doi:10.3389/fgene.2016.00019. PMC 4759636. PMID 26925095.
- ^ Miyawaki, A.; Llopis, J.; Heim, R.; McCaffery, J. M.; Adams, J. A.; Ikura, M.; Tsien, R. Y. (1997-08-28). "Fluorescent indicators for Ca2+ based on green fluorescent proteins and calmodulin". Nature. 388 (6645): 882–887. doi:10.1038/42264. ISSN 0028-0836. PMID 9278050.
- ^ Kerr, R.; Lev-Ram, V.; Baird, G.; Vincent, P.; Tsien, R. Y.; Schafer, W. R. (2000-06-01). "Optical imaging of calcium transients in neurons and pharyngeal muscle of C. elegans". Neuron. 26 (3): 583–594. doi:10.1016/s0896-6273(00)81196-4. ISSN 0896-6273. PMID 10896155.
- ^ Fiala, André; Spall, Thomas; Diegelmann, Sören; Eisermann, Beate; Sachse, Silke; Devaud, Jean-Marc; Buchner, Erich; Galizia, C. Giovanni (2002-10-29). "Genetically expressed cameleon in Drosophila melanogaster is used to visualize olfactory information in projection neurons". Current Biology. 12 (21): 1877–1884. doi:10.1016/s0960-9822(02)01239-3. ISSN 0960-9822. PMID 12419190.
- ^ Higashijima, Shin-ichi; Masino, Mark A.; Mandel, Gail; Fetcho, Joseph R. (2003-12-01). "Imaging neuronal activity during zebrafish behavior with a genetically encoded calcium indicator". Journal of Neurophysiology. 90 (6): 3986–3997. doi:10.1152/jn.00576.2003. ISSN 0022-3077. PMID 12930818.
- ^ Ji, Guangju; Feldman, Morris E.; Deng, Ke-Yu; Greene, Kai Su; Wilson, Jason; Lee, Jane C.; Johnston, Robyn C.; Rishniw, Mark; Tallini, Yvonne (2004-05-14). "Ca2+-sensing transgenic mice: postsynaptic signaling in smooth muscle". The Journal of Biological Chemistry. 279 (20): 21461–21468. doi:10.1074/jbc.M401084200. ISSN 0021-9258. PMID 14990564.
- ^ Nakai, J.; Ohkura, M.; Imoto, K. (2001-02-01). "A high signal-to-noise Ca(2+) probe composed of a single green fluorescent protein". Nature Biotechnology. 19 (2): 137–141. doi:10.1038/84397. ISSN 1087-0156. PMID 11175727.
- ^ Chen, Tsai-Wen; Wardill, Trevor J.; Sun, Yi; Pulver, Stefan R.; Renninger, Sabine L.; Baohan, Amy; Schreiter, Eric R.; Kerr, Rex A.; Orger, Michael B. (2013-07-18). "Ultrasensitive fluorescent proteins for imaging neuronal activity". Nature. 499 (7458): 295–300. Bibcode:2013Natur.499..295C. doi:10.1038/nature12354. ISSN 1476-4687. PMC 3777791. PMID 23868258.
- ^ "The Brain Prize 2013". Retrieved 3 October 2013.
- ^ Reiner, A.; Isacoff, E.Y. (October 2013). "The Brain Prize 2013: the optogenetics revolution". Trends Neurosci. 36 (10): 557–60. doi:10.1016/j.tins.2013.08.005. PMID 24054067.
- ^ a b c d Baratta M.V., Nakamura S, Dobelis P., Pomrenze M.B., Dolzani S.D. & Cooper D.C. (2012) Optogenetic control of genetically-targeted pyramidal neuron activity in prefrontal cortex. Nature Precedings April 2 doi=10.1038/npre.2012.7102.1 http://www.neuro-cloud.net/nature-precedings/baratta
- ^ Husson, S. J.; Liewald, J. F.; Schultheis, C.; Stirman, J. N.; Lu, H.; Gottschalk, A. (2012). Samuel, Aravinthan, ed. "Microbial Light-Activatable Proton Pumps as Neuronal Inhibitors to Functionally Dissect Neuronal Networks in C. Elegans". PLoS ONE. 7 (7): e40937. Bibcode:2012PLoSO...740937H. doi:10.1371/journal.pone.0040937. PMC 3397962. PMID 22815873.

- ^ Liu, Y.; Lebeouf, B.; Guo, X.; Correa, P. A.; Gualberto, D. G.; Lints, R.; Garcia, L. R. (2011). Goodman, Miriam B, ed. "A Cholinergic-Regulated Circuit Coordinates the Maintenance and Bi-Stable States of a Sensory-Motor Behavior during Caenorhabditis elegans Male Copulation". PLoS Genetics. 7 (3): e1001326. doi:10.1371/journal.pgen.1001326. PMC 3053324. PMID 21423722.
- ^ Akemann, W; Mutoh, H; Perron, A; Park, YK; Iwamoto, Y; Knöpfel, T (2012). "Imaging neural circuit dynamics with a voltage-sensitive fluorescent protein". J Neurophysiol. 108 (8): 2323–37. doi:10.1152/jn.00452.2012. PMID 22815406.
- ^ Deisseroth, Karl. "Optogenetics: Controlling the Brain with Light [Extended Version]". Scientific American. Retrieved 2016-11-28.
- ^ Beck, Sebastian; Yu-Strzelczyk, Jing; Pauls, Dennis; Constantin, Oana M.; Gee, Christine E.; Ehmann, Nadine; Kittel, Robert J.; Nagel, Georg; Gao, Shiqiang (2018). "Synthetic Light-Activated Ion Channels for Optogenetic Activation and Inhibition". Frontiers in Neuroscience. 12. doi:10.3389/fnins.2018.00643. ISSN 1662-453X.
- ^ Andrea Bernal Sierra, Yinth; Rost, Benjamin; Oldani, Silvia; Schneider-Warme, Franziska; Seifert, Reinhard; Schmitz, Dietmar; Hegemann, Peter (November 5, 2018). "Potassium channel-based two component optogenetic tool for silencing of excitable cells". Biophysical Journal. 114 (3): 668a. doi:10.1016/j.bpj.2017.11.3607. ISSN 0006-3495.
- ^ Zhao, S.; Cunha, C.; Zhang, F.; Liu, Q.; Gloss, B.; Deisseroth, K.; Augustine, G. J.; Feng, G. (2008). "Improved expression of halorhodopsin for light-induced silencing of neuronal activity". Brain Cell Biology. 36 (1–4): 141–154. doi:10.1007/s11068-008-9034-7. PMC 3057022. PMID 18931914.
- ^ Gradinaru, V.; Thompson, K. R.; Deisseroth, K. (2008). "ENpHR: A Natronomonas halorhodopsin enhanced for optogenetic applications". Brain Cell Biology. 36 (1–4): 129–139. doi:10.1007/s11068-008-9027-6. PMC 2588488. PMID 18677566.
- ^ a b c Witten, I. B.; Lin, S. C.; Brodsky, M.; Prakash, R.; Diester, I.; Anikeeva, P.; Gradinaru, V.; Ramakrishnan, C.; Deisseroth, K. (2010). "Cholinergic interneurons control local circuit activity and cocaine conditioning". Science. 330 (6011): 1677–81. Bibcode:2010Sci...330.1677W. doi:10.1126/science.1193771. PMC 3142356. PMID 21164015.
- ^ Kim, J. M.; Hwa, J.; Garriga, P.; Reeves, P. J.; RajBhandary, U. L.; Khorana, H. G. (2005). "Light-driven activation of beta 2-adrenergic receptor signaling by a chimeric rhodopsin containing the beta 2-adrenergic receptor cytoplasmic loops". Biochemistry. 44 (7): 2284–92. doi:10.1021/bi048328i. PMID 15709741.
- ^ Airan, R. D.; Thompson, K. R.; Fenno, L. E.; Bernstein, H.; Deisseroth, K. (2009). "Temporally precise in vivo control of intracellular signalling". Nature. 458 (7241): 1025–9. Bibcode:2009Natur.458.1025A. doi:10.1038/nature07926. PMID 19295515.
- ^ Levskaya, Anselm; Weiner, Orion D.; Lim, Wendell A.; Voigt, Christopher A. (October 2009). "Spatiotemporal control of cell signalling using a light-switchable protein interaction". Nature. 461 (7266): 997–1001. Bibcode:2009Natur.461..997L. doi:10.1038/nature08446. PMC 2989900. PMID 19749742.
- ^ Wu, Yi I.; Frey, Daniel; Lungu, Oana I.; Jaehrig, Angelika; Schlichting, Ilme; Kuhlman, Brian; Hahn, Klaus M. (September 2009). "A genetically encoded photoactivatable Rac controls the motility of living cells". Nature. 461 (7260): 104–8. Bibcode:2009Natur.461..104W. doi:10.1038/nature08241. PMC 2766670. PMID 19693014.
- ^ Yazawa, M.; Sadaghiani, A. M.; Hsueh, B.; Dolmetsch, R. E. (2009). "Induction of protein-protein interactions in live cells using light". Nature Biotechnology. 27 (10): 941–5. doi:10.1038/nbt.1569. PMID 19801976.
- ^ Stierl, M.; Stumpf, P.; Udwari, D.; Gueta, R.; Hagedorn, R.; Losi, A.; Gartner, W.; Petereit, L.; et al. (January 2011). "Light modulation of cellular cAMP by a small bacterial photoactivated adenylyl cyclase, bPAC, of the soil bacterium Beggiatoa". J. Biol. Chem. 286 (2): 1181–8. doi:10.1074/jbc.M110.185496. PMC 3020725. PMID 21030594.
- ^ Ryu, M.-H.; Moskvin, O. V.; Siltberg-Liberles, J.; Gomelsky, M. (December 2010). "Natural and engineered photoactivated nucleotidyl cyclases for optogenetic applications". J. Biol. Chem. 285 (53): 41501–8. doi:10.1074/jbc.M110.177600. PMC 3009876. PMID 21030591.
- ^ Lerner, TN; Ye, L; Deisseroth, K (2016). "Communication in Neural Circuits: Tools, Opportunities, and Challenges". Cell. 164 (6): 1136–50. doi:10.1016/j.cell.2016.02.027. PMID 26967281.
- ^ Aravanis, Alexander M; Wang, Li-Ping; Zhang, Feng; Meltzer, Leslie A; Mogri, Murtaza Z; Schneider, M Bret; Deisseroth, Karl (September 2007). "An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology". J Neural Eng. 4 (3): S143–56. Bibcode:2007JNEng...4S.143A. doi:10.1088/1741-2560/4/3/S02. PMID 17873414.. PMID 17873414
- ^ Adamantidis, Antoine R.; Zhang, Feng; Aravanis, Alexander M.; Deisseroth, Karl; De Lecea, Luis (November 2007). "Neural substrates of awakening probed with optogenetic control of hypocretin neurons". Nature. 450 (7168): 420–4. Bibcode:2007Natur.450..420A. doi:10.1038/nature06310. PMID 17943086.
- ^ a b Gradinaru, V.; Thompson, K. R.; Zhang, F.; Mogri, M.; Kay, K.; Schneider, M. B.; Deisseroth, K. (2007). "Targeting and readout strategies for fast optical neural control in vitro and in vivo". J. Neurosci. 27 (52): 14231–8. doi:10.1523/JNEUROSCI.3578-07.2007. PMID 18160630.
- ^ Damestani, Yasaman; Reynolds, Carissa L.; Szu, Jenny; Hsu, Mike S.; Kodera, Yasuhiro; Binder, Devin K.; Park, B. Hyle; Garay, Javier E.; Rao, Masaru P.; Aguilar, Guillermo (2013). "Transparent nanocrystalline yttria-stabilized-zirconia calvarium prosthesis". Nanomedicine. Elsevier Inc. 9 (8): 1135–8. doi:10.1016/j.nano.2013.08.002. PMID 23969102. Retrieved September 11, 2013. • Explained by Mohan, Geoffrey (September 4, 2013). "A window to the brain? It's here, says UC Riverside team". Los Angeles Times.
- ^ Wentz, Christian T; Bernstein, Jacob G; Monahan, Patrick; Guerra, Alexander; Rodriguez, Alex; Boyden, Edward S (2011). "A wirelessly powered and controlled device for optical neural control of freely-behaving animals". Journal of Neural Engineering. 8 (4): 046021. Bibcode:2011JNEng...8d6021W. doi:10.1088/1741-2560/8/4/046021. PMC 3151576. PMID 21701058.
- ^ Pama, E. A. Claudia; Colzato, Lorenza S.; Hommel, Bernhard (2013-01-01). "Optogenetics as a neuromodulation tool in cognitive neuroscience". Cognition. 4: 610. doi:10.3389/fpsyg.2013.00610. PMC 3764402. PMID 24046763.
- ^ Warden, Melissa R.; Cardin, Jessica A.; Deisseroth, Karl (2014-07-11). "Optical Neural Interfaces". Annual Review of Biomedical Engineering. 16: 103–129. doi:10.1146/annurev-bioeng-071813-104733. ISSN 1523-9829. PMC 4163158. PMID 25014785.
- ^ Guru, Akash; Post, Ryan J; Ho, Yi-Yun; Warden, Melissa R (2015-07-25). "Making Sense of Optogenetics". International Journal of Neuropsychopharmacology. 18 (11): pyv079. doi:10.1093/ijnp/pyv079. ISSN 1461-1457. PMC 4756725. PMID 26209858.
- ^ a b c d e Tischer, Doug; Weiner, Orion D. (2014-08-01). "Illuminating cell signalling with optogenetic tools". Nature Reviews Molecular Cell Biology. 15 (8): 551–558. doi:10.1038/nrm3837. ISSN 1471-0072. PMC 4145075. PMID 25027655.
- ^ a b c d e Zalocusky, Kelly A; Fenno, Lief E; Deisseroth, Karl (2013). "Current Challenges in Optogenetics". Society for Neuroscience.
- ^ Heitmann, Stewart; Rule, Michael; Truccolo, Wilson; Ermentrout, Bard (2017). "Optogenetic stimulation shifts the excitability of cerebral cortex from Type I to Type II: Oscillation onset and wave propagation". PLOS Computational Biology. 13 (1): e1005349. Bibcode:2017PLSCB..13E5349H. doi:10.1371/journal.pcbi.1005349. ISSN 1553-7358.
- ^ Lu, Yao; Truccolo, Wilson; Wagner, Fabien B.; Vargas-Irwin, Carlos E.; Ozden, Ilker; Zimmermann, Jonas B.; May, Travis; Agha, Naubahar S.; Wang, Jing; Nurmikko, Arto V. (2016). "Optogenetically induced spatiotemporal gamma oscillations and neuronal spiking activity in primate motor cortex". Journal of Neurophysiology. 113 (10): 3574–3587. doi:10.1152/jn.00792.2014. ISSN 1522-1598. PMC 4461886.
- ^ Gradinaru, Viviana; Thompson, Kimberly R.; Deisseroth, Karl (2008-08-01). "eNpHR: a Natronomonas halorhodopsin enhanced for optogenetic applications". Brain Cell Biology. 36 (1–4): 129–139. doi:10.1007/s11068-008-9027-6. ISSN 1559-7113. PMC 2588488. PMID 18677566.
- ^ a b Leergaard, Trygve B.; Hilgetag, Claus C.; Sporns, Olaf (2012-05-01). "Mapping the Connectome: Multi-Level Analysis of Brain Connectivity". Frontiers in Neuroinformatics. 6. doi:10.3389/fninf.2012.00014. ISSN 1662-5196. PMC 3340894. PMID 22557964.
- ^ Penzkofer A, Hegemann P, Kateriya S (2018). "Organic dyes in optogenetics". In Duarte FJ. Organic Lasers and Organic Photonics. London: Institute of Physics. pp. 13-1 to 13-114. ISBN 978-0-7503-1570-8.CS1 maint: Multiple names: authors list (link) CS1 maint: Uses editors parameter (link)
- ^ Kravitz, A. V.; Freeze, B. S.; Parker, P. R. L.; Kay, K.; Thwin, M. T.; Deisseroth, K.; Kreitzer, A. C. (2010). "Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry". Nature. 466 (7306): 622–626. Bibcode:2010Natur.466..622K. doi:10.1038/nature09159. PMC 3552484. PMID 20613723.
- ^ Gradinaru, V.; Mogri, M.; Thompson, K. R.; Henderson, J. M.; Deisseroth, K. (2009). "Optical Deconstruction of Parkinsonian Neural Circuitry". Science. 324 (5925): 354–359. Bibcode:2009Sci...324..354G. doi:10.1126/science.1167093. PMID 19299587.
- ^ Cardin, J. A.; Carlén, M.; Meletis, K.; Knoblich, Ulf; Zhang, Feng; Deisseroth, Karl; Tsai, Li-Huei; Moore, Christopher I. (2009). "(June 2009). "Driving fast-spiking cells induces gamma rhythm and controls sensory responses". Nature. 459 (7247): 663–7. Bibcode:2009Natur.459..663C. doi:10.1038/nature08002. PMC 3655711. PMID 19396156.
- ^ Sohal, V. S.; Zhang, F.; Yizhar, O.; Deisseroth, K. (2009). "Parvalbumin neurons and gamma rhythms enhance cortical circuit performance". Nature. 459 (7247): 698–702. Bibcode:2009Natur.459..698S. doi:10.1038/nature07991. PMC 3969859. PMID 19396159.
- ^ Tsai, H.C.; Zhang, F.; Adamantidis, A.; Stuber, G. D.; Bonci, A.; De Lecea, L.; Deisseroth, K. (2009). "Phasic firing in dopaminergic neurons is sufficient for behavioral conditioning". Science. 324 (5930): 1080–4. Bibcode:2009Sci...324.1080T. doi:10.1126/science.1168878. PMID 19389999.
- ^ Haubensak, W.; Kunwar, P. S.; Cai, H.; Ciocchi, S.; Wall, N. R.; Ponnusamy, R.; Biag, J.; Dong, H. W.; Deisseroth, K.; Callaway, E. M.; Fanselow, M. S.; Lüthi, A.; Anderson, D. J. (2010). "Genetic dissection of an amygdala microcircuit that gates conditioned fear". Nature. 468 (7321): 270–276. Bibcode:2010Natur.468..270H. doi:10.1038/nature09553. PMC 3597095. PMID 21068836.
- ^ Johansen, J. P.; Hamanaka, H.; Monfils, M. H.; Behnia, R.; Deisseroth, K.; Blair, H. T.; Ledoux, J. E. (2010). "Optical activation of lateral amygdala pyramidal cells instructs associative fear learning". Proceedings of the National Academy of Sciences. 107 (28): 12692–12697. Bibcode:2010PNAS..10712692J. doi:10.1073/pnas.1002418107. PMC 2906568. PMID 20615999.
- ^ Jasnow AM, Ehrlich DE, Choi DC, Dabrowska J, Bowers ME, McCullough KM, Rainnie DG, Ressler KJ (2013). "Thy1-expressing neurons in the basolateral amygdala may mediate fear inhibition". J. Neurosci. 33 (25): 10396–404. doi:10.1523/JNEUROSCI.5539-12.2013. PMC 3685835. PMID 23785152.
- ^ Dias, B. G.; Banerjee, S. B.; Goodman, J. V.; Ressler, K. J. (2013). "Towards new approaches to disorders of fear and anxiety". Current Opinion in Neurobiology. 23 (3): 346–352. doi:10.1016/j.conb.2013.01.013. PMC 3672317. PMID 23402950.
- ^ Karalis, Nikolaos; Dejean, Cyril; Chaudun, Fabrice; Khoder, Suzana; Rozeske, Robert R.; Wurtz, Hélène; Bagur, Sophie; Benchenane, Karim; Sirota, Anton (2016-04-01). "4-Hz oscillations synchronize prefrontal-amygdala circuits during fear behavior". Nature Neuroscience. 19 (4): 605–612. doi:10.1038/nn.4251. ISSN 1546-1726. PMC 4843971. PMID 26878674.
- ^ Shusterman, R; Smear, MC; Koulakov, AA; Rinberg, D (17 July 2011). "Precise olfactory responses tile the sniff cycle". Nature Neuroscience. 14 (8): 1039–44. doi:10.1038/nn.2877. PMID 21765422.
- ^ Smith RS, Hu R, DeSouza A, Eberly CL, Krahe K, Chan W, Araneda RC (Jul 2015). "Differential Muscarinic Modulation in the Olfactory Bulb". The Journal of Neuroscience. 35 (30): 10773–85. doi:10.1523/JNEUROSCI.0099-15.2015. PMC 4518052. PMID 26224860.
- ^ Patterson, Michael Andrew; Lagier, Samuel; Carleton, Alan (2013-08-27). "Odor representations in the olfactory bulb evolve after the first breath and persist as an odor afterimage". Proceedings of the National Academy of Sciences. 110 (35): E3340–E3349. Bibcode:2013PNAS..110E3340P. doi:10.1073/pnas.1303873110. ISSN 0027-8424. PMC 3761593. PMID 23918364.
- ^ Tecuapetla, F.; Patel, J. C.; Xenias, H.; English, D.; Tadros, I.; Shah, F.; Berlin, J.; Deisseroth, K.; Rice, M. E.; Tepper, J. M.; Koos, T. (2010). "Glutamatergic Signaling by Mesolimbic Dopamine Neurons in the Nucleus Accumbens". Journal of Neuroscience. 30 (20): 7105–7110. doi:10.1523/JNEUROSCI.0265-10.2010. PMC 3842465. PMID 20484653.
- ^ Bingen BO, Engels MC, Schalij MJ, Jangsangthong W, Neshati Z, Feola I, et al. (2014). "Light-induced termination of spiral wave arrhythmias by optogenetic engineering of atrial cardiomyocytes". Cardiovasc Res. 104 (1): 194–205. doi:10.1093/cvr/cvu179. PMID 25082848.
- ^ Nussinovitch, Udi; Gepstein, Lior (2015). "Optogenetics for in vivo cardiac pacing and resynchronization therapies". Nature Biotechnology. 33 (7): 750–754. doi:10.1038/nbt.3268. PMID 26098449.
- ^ Nyns EC, et al. (2016). "Optogenetic termination of ventricular arrhythmias in the whole heart: towards biological cardiac rhythm management". Eur Heart J: ehw574. doi:10.1093/eurheartj/ehw574.
- ^ Bruegmann T, et al. (2016). "Optogenetic defibrillation terminates ventricular arrhythmia in mouse hearts and human simulations". J Clin Invest. 126: 3894–3904. doi:10.1172/JCI88950. PMC 5096832.
- ^ Crocini C, et al. (2016). "Optogenetics design of mechanistically-based stimulation patterns for cardiac defibrillation". Sci Rep. 6. Bibcode:2016NatSR...635628C. doi:10.1038/srep35628.
- ^ Hernandez, Victor H.; et al. (2014). "Optogenetic stimulation of the auditory pathway". J Clin Invest. 124 (3): 1114–1129. doi:10.1172/JCI69050. PMC 3934189. PMID 24509078.
- ^ Mager T, Lopez de la Morena D, Senn V, Schlotte J, D'Errico A, Feldbauer K, Wrobel C, Jung S, Bodensiek K, Rankovic V, Browne L, Huet A, Jüttner J, Wood PG, Letzkus JJ, Moser T, Bamberg E (May 2018). "High frequency neural spiking and auditory signaling by ultrafast red-shifted optogenetics". Nat Commun. 9 (1): 1750. Bibcode:2018NatCo...9.1750M. doi:10.1038/s41467-018-04146-3. PMID 29717130.
- ^ Moser, Tobias (2015). "Optogenetic stimulation of the auditory pathway for research and future prosthetics". Current Opinion in Neurobiology. 34: 29–36. doi:10.1016/j.conb.2015.01.004. PMID 25637880.
- ^ Lin, JY; Knutsen, PM; Muller, A; Kleinfeld, D; Tsien, RY (2013). "ReaChR: A red-shifted variant of channelrhodopsin enables deep transcranial optogenetic excitation". Nature Neuroscience. 16 (10): 1499–1508. doi:10.1038/nn.3502. PMC 3793847. PMID 23995068.
- ^ Matthews, Gillian A.; Nieh, Edward H.; Vander Weele, Caitlin M.; Halbert, Sarah A.; Pradhan, Roma V.; Yosafat, Ariella S.; Glober, Gordon F.; Izadmehr, Ehsan M.; Thomas, Rain E. (2016-02-11). "Dorsal Raphe Dopamine Neurons Represent the Experience of Social Isolation". Cell. 164 (4): 617–631. doi:10.1016/j.cell.2015.12.040. ISSN 1097-4172. PMC 4752823. PMID 26871628.
- ^ Kovacs KA, O'Neill J, Schoenenberger P, Penttonen M, Ranguel Guerrero DK, Csicsvari J (19 Nov 2016). "Optogenetically Blocking Sharp Wave Ripple Events in Sleep Does Not Interfere with the Formation of Stable Spatial Representation in the CA1 Area of the Hippocampus". PLOS ONE. 11: e0164675. Bibcode:2016PLoSO..1164675K. doi:10.1371/journal.pone.0164675. PMC 5070819. PMID 27760158.
- ^ Valon, Léo; Marín-Llauradó, Ariadna; Wyatt, Thomas; Charras, Guillaume; Trepat, Xavier (2017-02-10). "Optogenetic control of cellular forces and mechanotransduction". Nature Communications. 8: 14396. Bibcode:2017NatCo...814396V. doi:10.1038/ncomms14396. ISSN 2041-1723. PMC 5309899. PMID 28186127.
- ^ Strickland, Devin; Lin, Yuan; Wagner, Elizabeth; Hope, C. Matthew; Zayner, Josiah; Antoniou, Chloe; Sosnick, Tobin R.; Weiss, Eric L.; Glotzer, Michael (2012-03-04). "TULIPs: tunable, light-controlled interacting protein tags for cell biology". Nature Methods. 9 (4): 379–384. doi:10.1038/nmeth.1904. ISSN 1548-7105. PMC 3444151. PMID 22388287.
- ^ Idevall-Hagren, Olof; Dickson, Eamonn J.; Hille, Bertil; Toomre, Derek K.; De Camilli, Pietro (2012-08-28). "Optogenetic control of phosphoinositide metabolism". Proceedings of the National Academy of Sciences of the United States of America. 109 (35): E2316–2323. Bibcode:2012PNAS..109E2316I. doi:10.1073/pnas.1211305109. ISSN 1091-6490. PMC 3435206. PMID 22847441.
- ^ Leung, Daisy W.; Otomo, Chinatsu; Chory, Joanne; Rosen, Michael K. (2008-09-02). "Genetically encoded photoswitching of actin assembly through the Cdc42-WASP-Arp2/3 complex pathway". Proceedings of the National Academy of Sciences of the United States of America. 105 (35): 12797–12802. Bibcode:2008PNAS..10512797L. doi:10.1073/pnas.0801232105. ISSN 1091-6490. PMC 2525560. PMID 18728185.
- ^ Toettcher, Jared E.; Gong, Delquin; Lim, Wendell A.; Weiner, Orion D. (2011-09-11). "Light-based feedback for controlling intracellular signaling dynamics". Nature Methods. 8 (10): 837–839. doi:10.1038/nmeth.1700. ISSN 1548-7105. PMC 3184382. PMID 21909100.
- ^ Lungu, Oana I.; Hallett, Ryan A.; Choi, Eun Jung; Aiken, Mary J.; Hahn, Klaus M.; Kuhlman, Brian (2012-04-20). "Designing photoswitchable peptides using the AsLOV2 domain". Chemistry & Biology. 19 (4): 507–517. doi:10.1016/j.chembiol.2012.02.006. ISSN 1879-1301. PMC 3334866. PMID 22520757.
- ^ Konermann, Silvana; Brigham, Mark D.; Trevino, Alexandro E.; Hsu, Patrick D.; Heidenreich, Matthias; Cong, Le; Platt, Randall J.; Scott, David A.; Church, George M. (2013-08-22). "Optical control of mammalian endogenous transcription and epigenetic states". Nature. 500 (7463): 472–476. Bibcode:2013Natur.500..472K. doi:10.1038/nature12466. ISSN 1476-4687. PMC 3856241. PMID 23877069.
- ^ Shimizu-Sato, Sae; Huq, Enamul; Tepperman, James M.; Quail, Peter H. (2002-10-01). "A light-switchable gene promoter system". Nature Biotechnology. 20 (10): 1041–1044. doi:10.1038/nbt734. ISSN 1087-0156. PMID 12219076.
- ^ Bugaj, Lukasz J.; Choksi, Atri T.; Mesuda, Colin K.; Kane, Ravi S.; Schaffer, David V. (2013-03-01). "Optogenetic protein clustering and signaling activation in mammalian cells". Nature Methods. 10 (3): 249–252. doi:10.1038/nmeth.2360. ISSN 1548-7105. PMID 23377377.
- ^ Zhou, Xin X.; Chung, Hokyung K.; Lam, Amy J.; Lin, Michael Z. (2012-11-09). "Optical control of protein activity by fluorescent protein domains". Science. 338 (6108): 810–814. Bibcode:2012Sci...338..810Z. doi:10.1126/science.1226854. ISSN 1095-9203. PMC 3702057. PMID 23139335.
- ^ Wu, Yi I.; Frey, Daniel; Lungu, Oana I.; Jaehrig, Angelika; Schlichting, Ilme; Kuhlman, Brian; Hahn, Klaus M. (2009-09-03). "A genetically encoded photoactivatable Rac controls the motility of living cells". Nature. 461 (7260): 104–108. Bibcode:2009Natur.461..104W. doi:10.1038/nature08241. ISSN 1476-4687. PMC 2766670. PMID 19693014.
- ^ Smart, Ashley D.; Pache, Roland A.; Thomsen, Nathan D.; Kortemme, Tanja; Davis, Graeme W.; Wells, James A. (2017-09-11). "Engineering a light-activated caspase-3 for precise ablation of neurons in vivo". Proceedings of the National Academy of Sciences of the United States of America. 114 (39): E8174–E8183. doi:10.1073/pnas.1705064114. ISSN 1091-6490. PMC 5625904. PMID 28893998.
- ^ Purvis, Jeremy E.; Lahav, Galit (2013-02-28). "Encoding and decoding cellular information through signaling dynamics". Cell. 152 (5): 945–956. doi:10.1016/j.cell.2013.02.005. ISSN 1097-4172. PMC 3707615. PMID 23452846.
- ^ Shimizu, Thomas S.; Tu, Yuhai; Berg, Howard C. (2010-06-22). "A modular gradient-sensing network for chemotaxis in Escherichia coli revealed by responses to time-varying stimuli". Molecular Systems Biology. 6: 382. doi:10.1038/msb.2010.37. ISSN 1744-4292. PMC 2913400. PMID 20571531.
- ^ Santos, Silvia D. M.; Verveer, Peter J.; Bastiaens, Philippe I. H. (2007-03-01). "Growth factor-induced MAPK network topology shapes Erk response determining PC-12 cell fate". Nature Cell Biology. 9 (3): 324–330. doi:10.1038/ncb1543. ISSN 1465-7392.
- ^ Toettcher, Jared E.; Weiner, Orion D.; Lim, Wendell A. (2013-12-05). "Using optogenetics to interrogate the dynamic control of signal transmission by the Ras/Erk module". Cell. 155 (6): 1422–1434. doi:10.1016/j.cell.2013.11.004. ISSN 1097-4172. PMC 3925772. PMID 24315106.
Further reading[edit]
This article's further reading may not follow Wikipedia's content policies or guidelines. Please improve this article by removing less relevant or redundant publications with the same point of view; or by incorporating the relevant publications into the body of the article through appropriate citations. (August 2014) (Learn how and when to remove this template message) |
- Airan, R. D.; Hu, E. S.; Vijaykumar, R.; Roy, M.; Meltzer, L. A.; Deisseroth, K. (October 2007). "Integration of light-controlled neuronal firing and fast circuit imaging". Current Opinion in Neurobiology. 17 (5): 587–92. doi:10.1016/j.conb.2007.11.003. PMID 18093822.
- Alilain, W. J.; Li, X.; Horn, K. P.; Dhingra, R.; et al. (November 2008). "Light-induced rescue of breathing after spinal cord injury". J. Neurosci. 28 (46): 11862–70. doi:10.1523/JNEUROSCI.3378-08.2008. PMC 2615537. PMID 19005051.
- Arenkiel, B. R.; Peca, J.; Davison, I. G.; Feliciano, Catia; et al. (April 2007). "In vivo light-induced activation of neural circuitry in transgenic mice expressing channelrhodopsin-2". Neuron. 54 (2): 205–18. doi:10.1016/j.neuron.2007.03.005. PMID 17442243.
- Atasoy, D.; Aponte, Y.; Su, H. H.; Sternson, S. M. (July 2008). "A FLEX switch targets Channelrhodopsin-2 to multiple cell types for imaging and long-range circuit mapping". J. Neurosci. 28 (28): 7025–30. doi:10.1523/JNEUROSCI.1954-08.2008. PMC 2593125. PMID 18614669.
- Ayling, O. G.; Harrison, T. C.; Boyd, J. D.; Goroshkov, A.; Murphy, T. H. (March 2009). "Automated light-based mapping of motor cortex by photoactivation of channelrhodopsin-2 transgenic mice". Nat. Methods. 6 (3): 219–24. doi:10.1038/nmeth.1303. PMID 19219033.
- Berndt, A.; Yizhar, O.; Gunaydin, L. A.; Hegemann, P.; Deisseroth, K. (February 2009). "Bi-stable neural state switches". Nature Neuroscience. 12 (2): 229–34. doi:10.1038/nn.2247. PMID 19079251.
- Bi, A.; Cui, J.; Ma, Y. P.; Olshevskaya, Elena; et al. (April 2006). "Ectopic expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration". Neuron. 50 (1): 23–33. doi:10.1016/j.neuron.2006.02.026. PMC 1459045. PMID 16600853.
- Busskamp, V.; Duebel, J.; Balya, D.; Fradot, M.; Viney, T. J.; Siegert, S.; Groner, A. C.; Cabuy, E.; Forster, V.; Seeliger, M.; Biel, M.; Humphries, P.; Paques, M.; Mohand-Said, S.; Trono, D.; Deisseroth, K.; Sahel, J. A.; Picaud, S.; Roska, B. (2010-07-23). "Genetic reactivation of cone photoreceptors restores visual responses in retinitis pigmentosa". Science. 329 (5990): 413–7. Bibcode:2010Sci...329..413B. doi:10.1126/science.1190897. PMID 20576849.
- Cardin, J. A.; Carlén, M.; Meletis, K.; Knoblich, U.; Zhang, F.; Deisseroth, K.; Tsai, L. H.; Moore, C. I. (2010). "Targeted optogenetic stimulation and recording of neurons in vivo using cell-type-specific expression of Channelrhodopsin-2". Nature Protocols. 5 (2): 247–54. doi:10.1038/nprot.2009.228. PMID 20134425.
- Carter, M. E.; Adamantidis, A.; Ohtsu, H.; Deisseroth, K.; de Lecea, L. (2009-09-02). "Sleep homeostasis modulates hypocretin-mediated sleep-to-wake transitions". Journal of Neuroscience. 29 (35): 10939–49. doi:10.1523/JNEUROSCI.1205-09.2009. PMC 3849591. PMID 19726652.
- Carter, M. E.; Yizhar, O.; Chikahisa, S.; Nguyen, H.; Adamantidis, A.; Nishino, S.; Deisseroth, K.; de Lecea, L. (December 2010). "Tuning arousal with optogenetic modulation of locus coeruleus neurons". Nature Neuroscience. 13 (12): 1526–33. doi:10.1038/nn.2682. PMC 3174240. PMID 21037585.
- Chow, B. Y.; Han, X.; Dobry, A. S.; Qian, Xiaofeng; et al. (January 2010). "High-performance genetically targetable optical neural silencing by light-driven proton pumps". Nature. 463 (7277): 98–102. Bibcode:2010Natur.463...98C. doi:10.1038/nature08652. PMC 2939492. PMID 20054397.
- Claridge-Chang, A.; Roorda, R. D.; Vrontou, E.; Sjulson, L.; Li, H.; Hirsh, J.; Miesenböck, G. (October 2009). "Writing memories with light-addressable reinforcement circuitry". Cell. 139 (2): 405–15. doi:10.1016/j.cell.2009.08.034. PMID 19837039.
- Clyne, J. D.; Miesenböck, G. (April 2008). "Sex-specific control and tuning of the pattern generator for courtship song in Drosophila". Cell. 133 (2): 354–63. doi:10.1016/j.cell.2008.01.050. PMID 18423205.
- Deisseroth, Karl. "Optogenetics: Controlling the Brain with Light".
- Diester, I.; Kaufman, M. T.; Mogri, M.; Pashaie, R.; Goo, W.; Yizhar, O.; Ramakrishnan, C.; Deisseroth, K.; Shenoy, K. V. (March 2011). "An optogenetic toolbox designed for primates". Nature Neuroscience. 14 (3): 387–97. doi:10.1038/nn.2749. PMC 3150193. PMID 21278729.
- Douglass, A. D.; Kraves, S.; Deisseroth, K.; Schier, A. F.; Engert, F. (August 2008). "Escape behavior elicited by single, channelrhodopsin-2-evoked spikes in zebrafish somatosensory neurons". Curr. Biol. 18 (15): 1133–7. doi:10.1016/j.cub.2008.06.077. PMC 2891506. PMID 18682213.
- Gradinaru, V.; Zhang, F.; Ramakrishnan, C.; Mattis, J.; Prakash, R.; Diester, I.; Goshen, I.; Thompson, K. R.; Deisseroth, K. (2010-04-02). "Molecular and cellular approaches for diversifying and extending optogenetics". Cell. 141 (1): 154–65. doi:10.1016/j.cell.2010.02.037. PMID 20303157.
- Gourine, A. V.; Kasymov, V.; Marina, N.; Tang, F.; Figueiredo, M. F.; Lane, S.; Teschemacher, A. G.; Spyer, K. M.; Deisseroth, K.; Kasparov, S. (2010-07-30). "Astrocytes control breathing through pH-dependent release of ATP". Science. 329 (5991): 571–5. Bibcode:2010Sci...329..571G. doi:10.1126/science.1190721. PMC 3160742. PMID 20647426.
- Gunaydin, L. A.; Yizhar, O.; Berndt, A.; Sohal, V. S.; Deisseroth, K.; Hegemann, P. (March 2010). "Ultrafast optogenetic control". Nature Neuroscience. 13 (3): 387–92. doi:10.1038/nn.2495. PMID 20081849.
- Han, X.; Boyden E. S.; Boyden (2007). Rustichini, Aldo, ed. "Multiple-color optical activation, silencing, and desynchronization of neural activity, with single-spike temporal resolution". PLoS ONE. 2 (3): e299. Bibcode:2007PLoSO...2..299H. doi:10.1371/journal.pone.0000299. PMC 1808431. PMID 17375185.CS1 maint: Multiple names: authors list (link)
- Han, X.; Qian, X.; Bernstein, J. G.; Zhou, Hui-hui; et al. (April 2009). "Millisecond-timescale optical control of neural dynamics in the nonhuman primate brain". Neuron. 62 (2): 191–8. doi:10.1016/j.neuron.2009.03.011. PMC 2830644. PMID 19409264.
- Hira, R.; Honkura, N.; Noguchi, J.; Maruyama, Yoshio; et al. (May 2009). "Transcranial optogenetic stimulation for functional mapping of the motor cortex". J. Neurosci. Methods. 179 (2): 258–63. doi:10.1016/j.jneumeth.2009.02.001. PMID 19428535.
- Hu, E. S.; Airan, R. D.; Vijaykumar, R.; Deisseroth, K. (July 2008). "Brain circuit dynamics". The American Journal of Psychiatry. 165 (7): 800. doi:10.1176/appi.ajp.2008.08050764. PMID 18593784.
- Huber, D.; Petreanu, L.; Ghitani, N.; Ranade, Sachin; et al. (January 2008). "Sparse optical microstimulation in barrel cortex drives learned behaviour in freely moving mice". Nature. 451 (7174): 61–4. Bibcode:2008Natur.451...61H. doi:10.1038/nature06445. PMC 3425380. PMID 18094685.
- Hwang, R. Y.; Zhong, L.; Xu, Y.; Johnson, T.; Zhang, F.; Deisseroth, K.; Tracey, W. D. (2007-12-18). "Nociceptive neurons protect Drosophila larvae from parasitoid wasps". Current Biology. 17 (24): 2105–16. doi:10.1016/j.cub.2007.11.029. PMC 2225350. PMID 18060782.
- Kuhlman, S. J.; Huang, Z. J.; Huang (2008). Wong, Rachel O. L., ed. "High-resolution labeling and functional manipulation of specific neuron types in mouse brain by Cre-activated viral gene expression". PLoS ONE. 3 (4): e2005. Bibcode:2008PLoSO...3.2005K. doi:10.1371/journal.pone.0002005. PMC 2289876. PMID 18414675.CS1 maint: Multiple names: authors list (link)
- Lagali, P. S.; Balya, D.; Awatramani, G. B.; Münch, Thomas A; et al. (June 2008). "Light-activated channels targeted to ON bipolar cells restore visual function in retinal degeneration". Nat. Neurosci. 11 (6): 667–75. doi:10.1038/nn.2117. PMID 18432197.
- Lee, J. H.; Durand, R.; Gradinaru, V.; Zhang, F.; Goshen, I.; Kim, D. S.; Fenno, L. E.; Ramakrishnan, C.; Deisseroth, K. (2010-06-10). "Global and local fMRI signals driven by neurons defined optogenetically by type and wiring". Nature. 465 (7299): 788–92. Bibcode:2010Natur.465..788L. doi:10.1038/nature09108. PMC 3177305. PMID 20473285.
- Li, H. H.; Roy, M.; Kuscuoglu, U.; Spencer, C. M.; Halm, B.; Harrison, K. C.; Bayle, J. H.; Splendore, A.; Ding, F.; Meltzer, L. A.; Wright, E.; Paylor, R.; Deisseroth, K.; Francke, U. (April 2009). "Induced chromosome deletions cause hypersociability and other features of Williams-Beuren syndrome in mice". EMBO Molecular Medicine. 1 (1): 50–65. doi:10.1002/emmm.200900003. PMC 3378107. PMID 20049703.
- Liewald, J. F.; Brauner, M.; Stephens, G. J.; Bouhours, Magali; et al. (October 2008). "Optogenetic analysis of synaptic function". Nat. Methods. 5 (10): 895–902. doi:10.1038/nmeth.1252. PMID 18794862.
- Lima, S. Q.; Hromádka, T.; Znamenskiy, P.; Zador, A. M.; Hromádka; Znamenskiy; Zador (2009). Nitabach, Michael N., ed. "PINP: a new method of tagging neuronal populations for identification during in vivo electrophysiological recording". PLoS ONE. 4 (7): e6099. Bibcode:2009PLoSO...4.6099L. doi:10.1371/journal.pone.0006099. PMC 2702752. PMID 19584920.CS1 maint: Multiple names: authors list (link)
- Lin, J. Y.; Lin, M. Z.; Steinbach, P.; Tsien, R. Y.; Lin; Steinbach; Tsien (March 2009). "Characterization of engineered channelrhodopsin variants with improved properties and kinetics". Biophys. J. 96 (5): 1803–14. Bibcode:2009BpJ....96.1803L. doi:10.1016/j.bpj.2008.11.034. PMC 2717302. PMID 19254539.CS1 maint: Multiple names: authors list (link)
- Liu, Q.; Hollopeter, G.; Jorgensen, E. M.; Hollopeter; Jorgensen (June 2009). "Graded synaptic transmission at the Caenorhabditis elegans neuromuscular junction". Proc. Natl. Acad. Sci. U.S.A. 106 (26): 10823–8. Bibcode:2009PNAS..10610823L. doi:10.1073/pnas.0903570106. PMC 2705609. PMID 19528650.CS1 maint: Multiple names: authors list (link)
- Llewellyn, M. E.; Thompson, K. R.; Deisseroth, K.; Delp, S. L. (October 2010). "Orderly recruitment of motor units under optical control in vivo". Nature Medicine. 16 (10): 1161–5. doi:10.1038/nm.2228. PMID 20871612.
- Lobo, M. K.; Covington, H. E., 3rd; Chaudhury, D.; Friedman, A. K.; Sun, H.; Damez-Werno, D.; Dietz, D. M.; Zaman, S.; Koo, J. W.; Kennedy, P. J.; Mouzon, E.; Mogri, M.; Neve, R. L.; Deisseroth, K.; Han, M. H.; Nestler, E. J. (2010-10-15). "Cell type-specific loss of BDNF signaling mimics optogenetic control of cocaine reward". Science. 330 (6002): 385–90. Bibcode:2010Sci...330..385L. doi:10.1126/science.1188472. PMC 3011229. PMID 20947769.CS1 maint: Multiple names: authors list (link)
- Miesenböck, G. (October 2008). "Lighting up the brain". Sci. Am. 299 (4): 52–9. Bibcode:2008SciAm.299d..52M. doi:10.1038/scientificamerican1008-52. PMID 18847085.
- Miesenböck, G. (October 2009). "The optogenetic catechism". Science. 326 (5951): 395–9. Bibcode:2009Sci...326..395M. doi:10.1126/science.1174520. PMID 19833960.
- Miller, G. (December 2006). "Optogenetics. Shining new light on neural circuits". Science. 314 (5806): 1674–6. doi:10.1126/science.314.5806.1674. PMID 17170269.
- Schneider, M. B.; Gradinaru, V.; Zhang, F.; Deisseroth, K. (May 2008). "Controlling neuronal activity". The American Journal of Psychiatry. 165 (5): 562. doi:10.1176/appi.ajp.2008.08030444. PMID 18450936.
- Schröder-Lang, S.; Schwärzel, M.; Seifert, R.; Strünker, Timo; et al. (January 2007). "Fast manipulation of cellular cAMP level by light in vivo". Nature Methods. 4 (1): 39–42. doi:10.1038/nmeth975. PMID 17128267.
- Szobota, S.; Gorostiza, P.; Del Bene, F.; Wyart, Claire; et al. (May 2007). "Remote control of neuronal activity with a light-gated glutamate receptor". Neuron. 54 (4): 535–45. doi:10.1016/j.neuron.2007.05.010. PMID 17521567.
- Toni, N.; Laplagne, D. A.; Zhao, C.; Lombardi, Gabriela; et al. (August 2008). "Neurons born in the adult dentate gyrus form functional synapses with target cells". Nat. Neurosci. 11 (8): 901–7. doi:10.1038/nn.2156. PMC 2572641. PMID 18622400.
- Tønnesen, J.; Sørensen, A. T.; Deisseroth, K.; Lundberg, C.; Kokaia, M. (2009-07-21). "Optogenetic control of epileptiform activity". Proceedings of the National Academy of Sciences of the United States of America. 106 (29): 12162–7. Bibcode:2009PNAS..10612162T. doi:10.1073/pnas.0901915106. PMC 2715517. PMID 19581573.
- Wang, S.; Szobota, S.; Wang, Y.; Volgraf, Matthew; et al. (December 2007). "All optical interface for parallel, remote, and spatiotemporal control of neuronal activity". Nano Lett. 7 (12): 3859–63. Bibcode:2007NanoL...7.3859W. doi:10.1021/nl072783t. PMID 18034506.
- Wang, H.; Peca, J.; Matsuzaki, M.; Matsuzaki, K.; et al. (May 2007). "High-speed mapping of synaptic connectivity using photostimulation in Channelrhodopsin-2 transgenic mice". Proc. Natl. Acad. Sci. U.S.A. 104 (19): 8143–8. Bibcode:2007PNAS..104.8143W. doi:10.1073/pnas.0700384104. PMC 1876585. PMID 17483470.
- Wang, Y.; Dye, C. A.; Sohal, V.; Long, J. E.; Estrada, R. C.; Roztocil, T.; Lufkin, T.; Deisseroth, K.; Baraban, S. C.; Rubenstein, J. L. (2010-04-14). "Dlx5 and Dlx6 regulate the development of parvalbumin-expressing cortical interneurons". Journal of Neuroscience. 30 (15): 5334–45. doi:10.1523/JNEUROSCI.5963-09.2010. PMC 2919857. PMID 20392955.
- Weick, J. P.; Johnson, M. A.; Skroch, S. P.; Williams, J. C.; Deisseroth, K.; Zhang, S. C. (November 2010). "Functional control of transplantable human ESC-derived neurons via optogenetic targeting". Stem Cells. 28 (11): 2008–16. doi:10.1002/stem.514. PMC 2988875. PMID 20827747.
- Zhang, F.; Wang, L. P.; Boyden, E. S.; Deisseroth, K. (October 2006). "Channelrhodopsin-2 and optical control of excitable cells". Nat. Methods. 3 (10): 785–92. doi:10.1038/nmeth936. PMID 16990810.
- Zhang, F.; Wang, L. P.; Brauner, M.; Liewald, Jana F.; et al. (April 2007). "Multimodal fast optical interrogation of neural circuitry". Nature. 446 (7136): 633–9. Bibcode:2007Natur.446..633Z. doi:10.1038/nature05744. PMID 17410168.
- Zhang, F.; Aravanis, A. M.; Adamantidis, A.; de Lecea, L.; Deisseroth, K. (August 2007). "Circuit-breakers: optical technologies for probing neural signals and systems". Nature Reviews Neuroscience. 8 (8): 577–81. doi:10.1038/nrn2192. PMID 17643087.
- Zhang, Y. P.; Holbro, N.; Oertner, T. G.; Holbro; Oertner (August 2008). "Optical induction of plasticity at single synapses reveals input-specific accumulation of alphaCaMKII". Proc. Natl. Acad. Sci. U.S.A. 105 (33): 12039–44. Bibcode:2008PNAS..10512039Z. doi:10.1073/pnas.0802940105. PMC 2575337. PMID 18697934.CS1 maint: Multiple names: authors list (link)
- Zhang, F.; Prigge, M.; Beyrière, F.; Tsunoda, Satoshi P; et al. (June 2008). "Red-shifted optogenetic excitation: a tool for fast neural control derived from Volvox carteri". Nat. Neurosci. 11 (6): 631–3. doi:10.1038/nn.2120. PMC 2692303. PMID 18432196.
- Zhang, F.; Gradinaru, V.; Adamantidis, A. R.; Durand, R.; Airan, R. D.; de Lecea, L.; Deisseroth, K. (2010). "Optogenetic interrogation of neural circuits: technology for probing mammalian brain structures". Nature Protocols. 5 (3): 439–56. doi:10.1038/nprot.2009.226. PMC 4503465. PMID 20203662.
- Zhang, J.; Laiwalla, F.; Kim, J. A.; Urabe, H.; Van Wagenen, R.; Song, Y. K.; Connors, B. W.; Zhang, F.; Deisseroth, K.; Nurmikko, A. V. (October 2009). "Integrated device for optical stimulation and spatiotemporal electrical recording of neural activity in light-sensitized brain tissue". Journal of neural engineering. 6 (5): 055007. Bibcode:2009JNEng...6e5007Z. doi:10.1088/1741-2560/6/5/055007. PMC 2921864. PMID 19721185.
- Zhu, P.; Narita, Y.; Bundschuh, S. T.; Fajardo, O.; Schärer, Y. P.; Chattopadhyaya, B.; Bouldoires, E. A.; Stepien, A. E.; Deisseroth, K.; Arber, S.; Sprengel, R.; Rijli, F. M.; Friedrich, R. W. (2009-12-11). "Optogenetic Dissection of Neuronal Circuits in Zebrafish using Viral Gene Transfer and the Tet System". Frontiers in Neural Circuits. 3: 21. doi:10.3389/neuro.04.021.2009. PMC 2805431. PMID 20126518.
- Zimmermann, G.; Wang, L. P.; Vaughan, A. G.; Manoli, D. S.; Zhang, F.; Deisseroth, K.; Baker, B. S.; Scott, M. P. (2009). Nitabach, Michael N., ed. "Manipulation of an innate escape response in Drosophila: photoexcitation of acj6 neurons induces the escape response". PLoS ONE. 4 (4): e5100. Bibcode:2009PLoSO...4.5100Z. doi:10.1371/journal.pone.0005100. PMC 2660433. PMID 19340304.
External links[edit]
This article's use of external links may not follow Wikipedia's policies or guidelines. (October 2013) (Learn how and when to remove this template message) |
- Optogenetics: shedding light on the brain's secrets Scientifica
- Optogenetics Resource Center, maintained by the Deisseroth lab.
- Synthetic Neurobiology Group, MIT, the portal of the Boyden lab.
- OpenOptogenetics.org, an optogenetics wiki, and its companion blog.
- Molecular Neurogenetics and Optophysiology Laboratory,"Optogenetic activation and silencing recordings of individual prefrontal cortical neurons in vivo and in vitro.
- Sohal lab portal
- Nurmikko lab portal
- Lab of Dr. Zhuo-Hua Pan
- Optophysiology at the Tyler lab
- Video: Ed Boyden on Optogenetics -- selective brain stimulation with light (SPIE Newsroom, April 2011)
| Technologies |  | |
|---|---|---|
| Scientific phenomena | ||
| Disciplines | ||
| Speculative | ||
| People | ||
| Other | ||